|
刷り込み行動を引きおこすには、親鳥の姿と声という目(視覚)と耳(聴覚)からの情報が重要です。
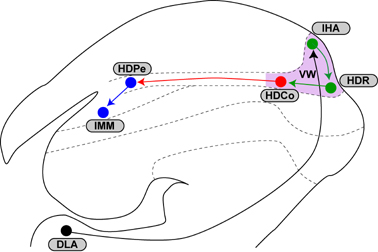
鳥類の視覚中枢の1つは、終脳のVW(visual Wulst;哺乳類の皮質視覚野に相当)と呼ばれる領域です。私たちの研究により、ここを破壊すると視覚刺激に対する刷り込みが起こらなくなること、さらに刷り込み学習によって、刷り込み刺激に対してVW内の反応する領域が変化することがわかりました(Maekawa et al., 2007)。一方で、刷り込みの記憶の貯蔵には、終脳のIMM(intermediate medial mesopallium;哺乳類の皮質連合野に相当)という脳領域が関与することがすでに知られていました。
そこで私たちは、目から入った情報がどのように処理されて、刷り込みが起こるのかを調べました。まず、VWからIMMへ至る神経伝達経路があるのかどうか、あるとしたらどのようなものか、に焦点をあてました。
1.VWからIMMへ至る神経経路の発見
順行性神経トレーサーのBDA(biotinylated dextran amine)及び逆行性神経トレーサーのCTb(cholera toxin subunit B) を用いて組織学的解析を行いました。その結果、①VWの腹側部のHDCo(core
region of the hyperpallium densocellulare)領域にある細胞は、IMMの背側にあるHDPe(periventricular part of the HD)領域へ長い軸索をのばしていること、②HDPe領域にある細胞は、その軸索をIMMへ投射していること、の2点が明らかになりました。また、急性脳スライスを用いた神経活動の光学的イメージング法により、VW→HDCo→IMMという経路(VW-IMM回路)を通ってシグナルが伝達されることが確認できました。さらに、薬理学的実験により、HDCo領域とHDPe領域では、主要な興奮性神経伝達物質であるグルタミン酸の受容体を介したシナプス伝達が起きていることがわかりました。
イボテン酸をHDCo領域に微量注入し神経細胞を破壊すると、刷り込みの成立が阻害されたため、刷り込みの成立にはHDCo細胞の活動が必須であることも示されました。
では、このVW-IMM回路は、刷り込みの成立にはどのように働いているのでしょうか。
2. VW-IMM回路の活動性の変化
A) 刷り込み学習成立の臨界期とVW-IMM回路の神経活動性の変化との関係
ヒナは大きくなってしまうと、刷り込まれなくなります。刷り込むことができる期間のことを、刷り込みの臨界期(あるいは感受性期)と呼びます。私たちの実験系では、これは孵化後1日目から4日目(P1からP4)の間です。光学的イメージングによってVW-IMM回路の神経活動性を調べると、臨界期中には電気的刺激に対する高い神経応答性が見られた一方で、臨界期終了後には応答性は著しく低下していました。これは、臨界期中にVW-IMM回路の神経活動性が高いことが刷り込み学習が成立する(刷り込まれる)ための条件であることを示唆していました。
B) 刷り込み学習の成立とVW-IMM回路の活動性の変化
臨界期中のP1に刷り込まれた個体は、臨界期終了後であるP7でもその図形を追いかける刷り込み行動を示しました。刷り込まれなかった個体(非刷り込み群)とは違って、これらの個体の脳では、VWからIMMへ至る強いシグナル伝達が観察されました。また、強く刷り込まれているほど(刷り込み学習の成績が良い)、強いシグナル伝達がみられました。このことから、臨界期終了以降も刷り込み群でみられるVW-IMM回路の高い神経活動性は、刷り込み記憶の保持に関わっていることが推測されました。
C) HDCo細胞の活性化と刷り込み
HDPe領域にCTbを注入して、HDCo細胞を逆行性に標識することで可視化できます。そして、HDCo細胞の活動性が刷り込みによってどのように変化するかについて、c-fosの発現を神経活動性の指標にして解析しました。その結果、刷り込み学習によりHDCo細胞、さらにHDCoから投射を受けるHDPeやIMMの細胞で活動性が上昇していました。このことから、VW-IMM回路の賦活化はHDCo細胞の活性化が引き金となっておきていることが示唆されました。
刷り込みによってHDCo細胞の活性化がおき、VW-IMM回路が賦活化されることがわかりましたが、HDCo細胞の活性化はどのようなメカニズムでおきているのでしょうか。
3.刷り込みによって起こる神経活動性変化のメカニズム
A)VW-IMM回路における NMDARの時期特異的、学習依存的な発現
グルタミン酸受容体の1つであるNMDA受容体(NMDAR)は、NR1と複数のNR2サブユニットから構成されるイオンチャネル型受容体です。組み合わされるNR2サブユニットの種類によって、NMDARの機能特性が異なり、また、生後の発達段階や組織によって、使われるNR2サブユニットが異なっていることがマウスやラットでは知られています。そこでNMDARのサブユニットを構成するNR1、NR2A、NR2BのmRNAの発現変化をin situ hybridizationによって調べたところ、P1ではNR2Aに比べNR2Bの発現が高いのですが、刷り込みを行うと、さらにNR1とNR2Bの発現細胞数が増加していました。また、臨界期終了後のP7ではNR2Bは減少し、代わりにNR2Aが増加していました。
B)NMDARの活性化と刷り込み成立の関係
HDCo細胞のNMDARやNR2Bの働きを阻害剤によって抑えておくと、刷り込まれなくなり、VW-IMM回路の神経活動も上昇しません。これは、刷り込みの成立にはHDCoにおけるNR2Bサブユニットを含むNMDARの活性化が必要であることを示しています。NR2Bを含むNMDARが活性化されると細胞内へのナトリウムイオン等の流入が長く続くため、シナプスが強化されやすいことが知られています。HDCo領域におけるNR2Bの発現が臨界期中に高く、さらに学習の刺激により増加することが、VW-IMM回路の神経活動性を高め、刷り込み学習の成立や維持に役立っていると考えられました。
4.おわりに
鳥類だけでなく多くの生物の生存にとって重要であると考えられるこの刷り込みは、速やかに起こります。本研究では、NR2B/NR1受容体の活動によって神経の活動性が高まることで、迅速な神経の可塑的変化が実現されていることを示すことができました。NR2Bの発現を学習に合わせて上昇させれば、臨界期中にみられるような高い可塑性を大人でも再現できるかもしれません。NR2Bの学習における役割の解明が進めば、NR2Bを標的とした薬剤の研究などを通じて、認知症やアルツハイマーなどの疾患の治療に役立つことが期待されます。 |

